转基因玉米HGK60在不同遗传背景下抗虫性鉴定及农艺性状分析
李鹏程 张明俊 王银晓 李香银 李圣彦 郎志宏
(中国农业科学院生物技术研究所,北京 100081)
玉米是重要的粮食作物、饲料和工业原料,种植面积与产量居三大农作物之首。虫害是造成玉米产量损失的重要因素,近年来全国玉米病虫害发生总体偏重,亚洲玉米螟、草地贪夜蛾、棉铃虫、黏虫是危害玉米的主要害虫,在局部地区造成玉米产量重大损失[1]。目前我国防控玉米害虫主要采用化学杀虫剂,但化学杀虫剂的大量使用伴随着一系列的环境污染、害虫抗药性增加等生态问题。随着转基因技术的发展,转Bt(Bacillus thuringiensis)基因抗虫玉米对有效防治玉米害虫具有重要作用。
自1996年转基因作物商业化种植以来,转基因作物种植面积增长迅速,2021年全球共有24 个国家种植了1.955 亿hm2的转基因作物,比2020年增加了3.7%(https://gm.agbioinvestor.com/)。转基因玉米种植面积在6 000 万hm2以上,仅次于转基因大豆,其主要性状为抗虫和耐除草剂[2]。统计结果表明种植抗虫转基因玉米可显著增加玉米产量和经济效益,据估算如果抗虫转基因玉米在我国商业化种植,GDP 可增加约86 亿美元,玉米自给率可增加约2%[3]。目前抗虫转基因玉米选用的杀虫基因主要为苏云金芽胞杆菌的cry类基因和vip类基因,其中防治鳞翅目害虫的cry1Ab基因是应用最广泛的杀虫基因,例如先正达公司的Bt11、孟山都公司的MON810、大北农公司的DBN9936 均含有cry1Ab基因,瑞丰125 含有cry1Ab与cry2Aj结构域融合基因,瑞丰8 和ND207 含有cry1Ab与cry2Ab双基因[4-5]。
我国在Bt 杀虫基因的发掘和克隆等方面发展迅速,取得了较好的进展,为抗虫转基因玉米的研发提供了基因资源。Btcry1Ah基因是从苏云金芽胞杆菌BT8 菌株中克隆获得的,与cry1Ab基因同属于cry1A家族,对亚洲玉米螟、棉铃虫、二化螟和小菜蛾等鳞翅目害虫具有高毒性[6-7]。在前期工作中将cry1Ah基因转入玉米,通过对转基因玉米转化群体进行抗虫性筛选,获得对玉米螟高抗的转化事件HGK60,对主要的玉米害虫亚洲玉米螟和棉铃虫具有高杀虫活性[8-10]。
Bt 杀虫基因在植物转化体中是否能够正常的表达,以及在不同世代、不同回交转育品系中能否稳定表达受到多方面的影响,比如外源基因启动子的选择、基因在玉米基因组中插入位置、密码子偏好性选择、回交受体的遗传背景以及不同的发育时期等[11]。前人研究表明,不同的遗传背景条件下,Bt杀虫蛋白的表达量会有不同,蛋白表达量和抗虫性正相关[12-14]。因此研究Bt 基因在不同遗传背景下的表达及抗虫稳定性,对转Bt 抗虫基因作物的应用推广具有重要意义。
本研究利用转Btcry1Ah基因玉米HGK60 回交转育获得的4 个自交系(HGK60-郑58、HGK60-昌7-2、HGK60-lx03-2、HGK60-lx05-4)和杂交组合HGK60-郑单958(HGK60-郑58 × HGK60-昌7-2)和HGK60- 鲁 单9066(HGK60-lx05-4 × HGK60-lx03-2),对其进行DNA 分子水平检测、外源蛋白含量检测、抗虫性鉴定以及农艺性状调查,分析HGK60 的遗传和抗虫稳定性,为我国生物育种工作提供一定的指导。
1.1 材料
1.1.1 玉米材料 转Btcry1Ah基因抗虫玉米自交系HGK60-郑58(BC6S3、BC6S4、BC6S5)、HGK60-昌7-2(BC6S3、BC6S4、BC6S5)、HGK60-lx03-2(BC6S3、BC6S4、BC6S5)、HGK60-lx05-4(BC6S3、BC6S4、BC6S5),玉米杂交种HGK60-郑单958(HGK60-郑58 × HGK60-昌7-2) 和HGK60- 鲁单9066(HGK60-lx05-4 ×HGK60-lx03-2)及其玉米自交系郑58 和昌7-2 由本实验室保存,玉米自交系lx03-2 和lx05-4 由山东省农业科学院玉米研究所孟昭东课题组提供。
1.1.2 试虫材料 亚洲玉米螟虫卵购自北京美延农业科技有限公司。虫卵置于人工培养箱(28℃)孵化。所有初孵幼虫作为供试虫源进行生物活性测定。
1.2 方法
1.2.1 试验设计 试验分小区种植,设置为3 次重复,每小区种植4 种转基因玉米自交系以及2 种杂交组合及其对应的非转基因对照玉米,共12 种材料,每个材料种植20 行,行长为5 m,行距为0.6 m,株距0.25 m。
1.2.2 转化体特异性PCR 鉴定 将供试玉米材料种植于大田,待玉米生长至V4 期时取4 种转基因自交系(HGK60-郑58、HGK60-昌7-2H、HGK60-lx03-2、HGK60-lx05-4)和2 种杂交组合(HGK60-郑单958 和HGK60-鲁单9066)以及非转基因对照郑95 叶片速冻在液氮中,采用CTAB 法提取基因组DNA,选用转化体特异性引物,引物序列为F: 5"-CGAAACAAGAAAAATGTTTCCTTAG-3" 和R:5"-GTGCGCGCTAGCTGGCGTGTT-3",进行PCR 扩增,扩增产物长度为628 bp,扩增条件为:94℃ 5 min;
94℃ 20 s,54℃ 20 s,72℃ 50 s,35 个循环;
72℃5 min。
1.2.3 ELISA 检测 选取种植于大田的转基因玉米及受体玉米V6 期叶片,液氮保存。植物总蛋白的提取以及Cry1Ah 蛋白含量检测,使用的是Cry1Ab/Cry1Ac ELISA 检测试剂盒(AP 003,Envirologix,USA),将100 mg 玉米叶片研磨成粉末转移至2.0 mL 离心管中,加入500 μL Extraction buffer 涡旋混匀,后续ELISA 步骤按照试剂盒说明书要求进行。Cry1Ah 标准蛋白浓度为1 000 ppb,制作标准曲线的推荐浓度为50、25、10、5、2、1 和0.5 ppb。
1.2.4 室内抗虫性鉴定 亚洲玉米螟的室内抗虫性鉴定参照刘晓贝等[14]的方法。玉米植株生长至V6期时,选取植株幼嫩心叶,用剪刀剪成1 cm2大小后放入24 孔接虫板中,然后用毛笔轻轻挑起玉米螟初孵幼虫放到接虫板中,每孔接一头幼虫,每个材料3 次生物学重复,从第2 天开始观察玉米螟初孵幼虫的存活情况,连续观察一周,并做好记录和拍照。
1.2.5 田间抗虫性鉴定 转基因玉米生长至小喇叭口期时,将60 头亚洲玉米螟初孵幼虫接到玉米心叶中,非转基因玉米作为阴性对照,3 d 后再接虫一次,接虫后第14 天和第21 天进行调查玉米被害情况。试验设计、接虫方法、调查方法、结果分析等按照《农业部953 号公告-10.1-2007》要求开展。
1.2.6 转基因玉米杂交组合农艺性状鉴定 分别在河北廊坊中国农业科学院国际农业高新技术产业园以及海南三亚中国农业科学院生物技术研究所南繁基地种植配制好的杂交组合HGK60-郑单958 和HGK60-鲁单9066 以及非转基因对照玉米。采用常规的田间管理方式,不进行任何害虫防治措施,除草、水肥按正常田间管理方式进行,待玉米收获前测量株高穗位高,收获后每个小区每个材料随机选取40个玉米穗进行穗长、穗粗、秃尖长、穗行数、行粒数、百粒重等农艺性状测量。使用SPSS 软件对数据进行统计分析。
2.1 转基因玉米特异性PCR鉴定
对回交获得的4 个转基因自交系(HGK60-郑58、HGK60- 昌7-2、HGK60-lx03-2、HGK60-lx05-4)、2 个杂交种HGK60-郑单958 和HGK60-鲁单9066 以及非转基因对照郑58 提取玉米基因组DNA,利用转化体特异性扩增引物进行PCR 分析,以质粒pm4AhG2 为阳性对照,郑58 玉米基因组DNA 为阴性对照,ddH2O 为空白对照,检测在不同的遗传背景中外源基因cry1Ah是否稳定插入,结果如图1所示,在4 个转基因自交系和2 个杂交种玉米中均能扩增出628 bp 的目标条带,证明通过回交转育的方式,Btcry1Ah抗虫基因已经成功转入到不同的玉米品种中。

图1 利用转化体特异性PCR 鉴定4 个转基因自交系和2个杂交组合Fig.1 Detection of 4 transgenic inbred lines and 2 hybrid combinations using event-special PCR
2.2 不同遗传背景转基因玉米中Cry1Ah蛋白表达量
取大田生长至V6 期的转基因玉米叶片通过ELISA检测不同遗传背景玉米中Cry1Ah蛋白表达量。结果显示在4 个转基因自交系和2 个杂交组合中,目的蛋白Cry1Ah 均有表达,而对照材料郑58 中无目的蛋白表达。其中,在自交系HGK60-lx03-2 中目的蛋白的表达量最高,平均值达到了3 528.9 ng/g FW(鲜重),自交系HGK60-lx05-4 中目的蛋白的表达量最低,平均值为2 895.8 ng/g FW。研究表明在不同自交系或者杂交种中,Cry1Ah 蛋白表达量无显著差异,cry1Ah基因在转基因玉米中已连续遗传10代以上,其表达量与低世代转化体相比,Cry1Ah 蛋白的表达量没有显著变化[9]。

图2 不同遗传背景转基因玉米叶片中Cry1Ah 蛋白表达分析Fig.2 Expression analysis of Cry1Ah protein in the transgenic maize leaves with different genetic backgrounds
2.3 转基因玉米室内玉米螟抗虫性鉴定
为了进一步检测HGK60 转基因玉米的抗虫性,分别在海南三亚和河北廊坊两年连续三代种植转基因玉米,生长至V6 叶期时,取玉米心叶接虫后进行室内玉米螟抗虫性鉴定,统计玉米螟死亡情况。连续3 代的玉米螟生测结果显示HGK60 的4 个转基因自交系和2 个转基因杂交组合中,接虫第2 天开始玉米螟幼虫出现死亡,供试叶片HGK60 有微小虫孔,接虫第3 天,不同遗传背景的HGK60 玉米已经表现出高杀虫活性,在2020年海南和2021年廊坊两地死亡率达到了100%,在2021年海南的死亡率到90%以上,个别试虫存活,但活力下降,接虫第4 天,供试玉米螟死亡率达100%,而非转基因对照玉米上玉米螟死亡率低于10%(表1)。
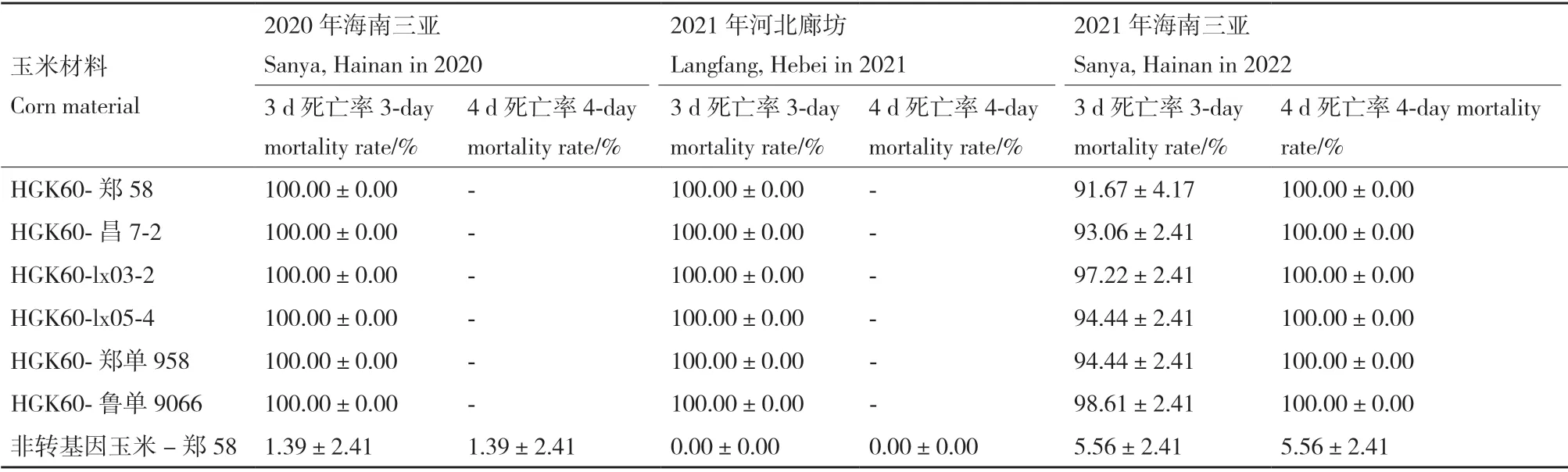
表1 亚洲玉米螟室内生测结果Table 1 Results of laboratory bioassay of Asian corn borer(Ostrinia furnacalis)
2.4 转基因玉米田间玉米螟抗虫性鉴定
玉米螟室内生测的同时,我们也在海南三亚和河北廊坊两地连续三代进行了转基因玉米田间玉米螟抗虫性鉴定。在小喇叭口期向玉米心叶接玉米螟,每株玉米接虫60 头左右,第14 天和第21 天后观察玉米叶片咬食情况,心叶抗虫级别鉴定依据《农业部953 号公告-10.1-2007》中9 级分级标准,1 为高抗,9 为高感。结果表明4 种转基因自交系及其配制的2种转基因杂交种对玉米螟的抗性等级均在1-2 之间,为高抗级别,非转基因对照则在6 级和7 级,为感虫级别(表2)。证明田间4 个转基因自交系以及杂交种HGK60-郑单958 和HGK60-鲁单9066 对玉米螟高抗。

表2 小喇叭口期亚洲玉米螟的食叶级别Table 2 Leaf feeding grades of the Asian corn borer in the maize small trumpet stage
2.5 转基因玉米杂交组合农艺性状
为了鉴定转基因玉米在农艺性状上的是否与非转基因对照有差异,连续三代对转基因及非转基因玉米株高穗位高以及其他产量相关性状进行统计分析,结果表明同一时期和地点的转基因玉米杂交种HGK60-郑单958 和HGK60-鲁单9066 与非转基因对照玉米比较时,株高、穗位高、穗长、穗粗、穗行数、行粒数、秃尖长、百粒重、单株产量上均无显著差异(表3),同时在不同的地点(河北廊坊和海南三亚)之间农艺性状表现出差异,造成该结果的主要原因是两地的自然条件存在明显差异,通过连续三代的农艺性状分析说明转基因玉米与非转基因对照玉米相比,两者之间农艺性状没有显著差异,外源基因的插入未对玉米受体材料农艺性状产生影响。
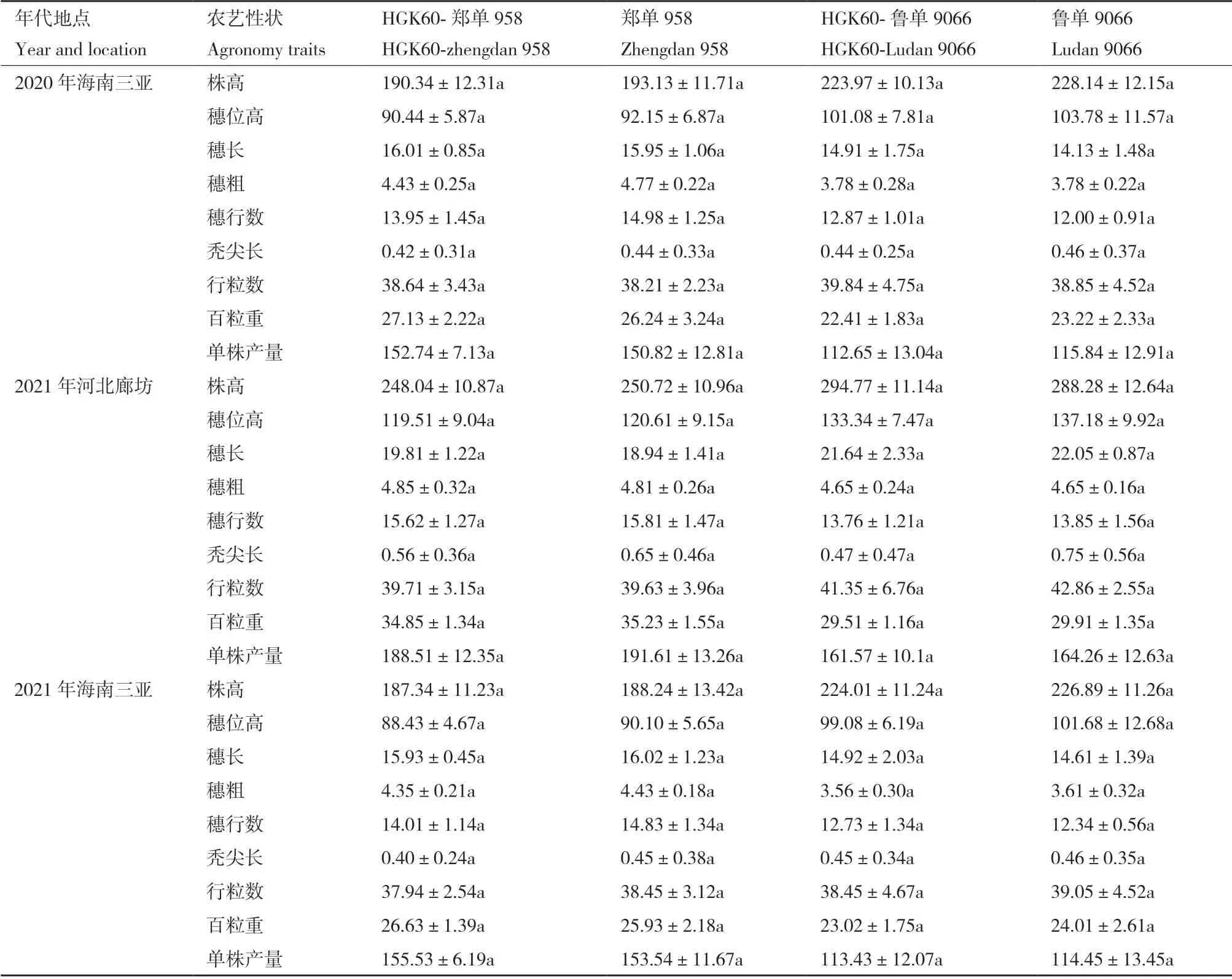
表3 转基因玉米杂交组合农艺性状分析Table 3 Agronomic traits analysis of transgenic maize hybrid lines
世界农业育种技术的发展主要经历了原始育种阶段、传统育种阶段及分子育种阶段[15]。转基因育种属于第一代分子育种技术,是改变农业生产方式最大的生物育种技术。2019-2022年我国已经批准了11 个抗虫、耐除草剂转基因玉米的生产应用安全证书,产业化试点工作正稳步推进中。在转基因玉米产业化应用中转化体与回交转育的玉米品种同等重要,在不同的玉米品种中外源基因是否可以稳定遗传、目标性状是否稳定、是否对玉米的农艺性状产生非预期影响等,这些问题的回答既需要针对不同转化体和受体品种一一对应来观察,也需要积累更多的研究数据,形成一些共性的结论,为转基因玉米产业化稳步推进和安全应用打好基础。
回交转育是将转基因性状导入商业化品种中所采用的最主要的方法。Venkatesh 等[16]将3 个不同性状转基因玉米(抗旱玉米MON 87460、耐草甘膦玉米NK603、抗虫玉米MON 89034)分别回交转育至6 个玉米骨干自交系,回交6 代后的转育材料与受体亲本基因型相似性理论值为99%,通过全基因组序列分析,不同回交转育的自交系与受体亲本基因型相似性在93.7%-99.35%之间。这表明通过多代的回交转育可使转育材料与亲本基因型基本达到一致。在本研究中抗虫玉米HGK60 已回交转育自交系6 代并自交3 代,表型和基因型与亲本材料趋于相同。通过转化体特异性PCR 检测,证明抗虫基因已稳定整合至玉米基因组中,4 个转基因自交系及2 个杂交种与回交受体遗传背景相同,具有转基因性状,是研究不同遗传背景对目标性状影响的合适材料。
在转基因育种过程中,外源基因插入到受体基因组上之后,可能会发生基因重组、沉默甚至丢失,导致外源蛋白在不同受体品种中不能表达或表达量低从而影响转基因性状的表现[17],因此选择表达量高的受体品种是首要任务[18]。本研究选用的HGK60 转基因玉米含有Bt 杀虫基因cry1Ah,ELISA研究表明在不同玉米受体品种中Cry1Ah 蛋白表达量没有显著差异,六叶期叶片中Cry1Ah 蛋白表达为2 895.8 ng/g-3 528.9 μg/g,与宋苗等[9]在T2代检测结果一致,说明cry1Ah基因在转基因玉米中稳定遗传和稳定表达。基于Cry1Ah 蛋白的稳定表达,室内抗虫性鉴定结果说明,第3 天时玉米螟初孵幼虫的死亡率达到100%,证明HGK60 转基因玉米在不同遗传背景下的杀虫活性是稳定的。Bt 蛋白在不同遗传背景玉米中表达量存在波动情况,如王培等[19]采用ELISA 方法研究在不同遗传背景下Cry1Ac 蛋白的表达量存在差异,波动范围为44.07-438.00 ng/g FW,但不影响杀虫效果。本研究导入4 个自交系和2 个杂交种中Cry1Ah 蛋白表达量没有比较大的波动,抗虫性稳定。
本实验室前期分别在海南三亚和河北廊坊两地对转HGK60 玉米郑单958H 与非转基因对照材料郑单958 进行接虫处理后测量株高、穗位高、穗长、穗粗、单株产量等农艺性状,结果显示转HGK60 玉米郑单958H 的单株产量明显高于非转基因对照材料郑单958,其余农艺性状除株高郑单958H 要相对高一些外无显著差异[10]。在本研究中转基因杂交种HGK60-郑单958 和HGK60-鲁单9066 与非转基因对照材料相比,株高和穗位高以及果穗的穗长、穗粗、行粒数、粒行数、秃尖长等相比无显著差异,说明cry1Ah基因的导入没有影响玉米的农艺性状,但在3 地环境中HGK60-鲁单9066 的百粒重和单株产量稍低于鲁单9066 的百粒重和单株产量,差异不显著,在下一步工作中需要增加试验点和加大试验群体来明确两者是否存在差异以及差异产生的原因。
HGK60 玉米具有抗虫性状,对于虫害发生不严重年份,转基因玉米与非转基因玉米的产量没有显著差异,但在虫害发生严重时,非转基因玉米的穗部受害虫危害严重,产量会显著降低。本研究结果表明,在郑单958 和鲁单9066 背景下HGK60 转基因玉米不会影响受体材料的农艺性状,并不能代表HGK60 转育到其他受体材料中仍然会有同样的结果,所以对于后续其他玉米品种的回交转育还是需要进行评估。随着转基因抗虫玉米产业化临近,优异抗虫转化体需要导入更多的骨干自交系和杂交种中,在转育过程中需重点关注外源基因遗传稳定性、抗虫效果及对农艺性状的影响,使优异转化体和优秀品种结合,保证转基因玉米产业化稳步推进。
以转Btcry1Ah基因的转基因玉米HGK60 为供体,将cry1Ah基因分别导入4 个玉米自交系并杂交获得2 个杂交种,cry1Ah基因在不同遗传背景玉米中稳定遗传和表达;
田间和室内玉米螟抗虫性鉴定表明不同遗传背景的转基因玉米对玉米螟高抗;
不同遗传背景转基因玉米与其受体对照玉米相比,两者之间农艺性状没有显著差异。