冷藏菲律宾蛤仔活体中优势菌的分离鉴定、生长特性与致腐能力分析
杨 溢, 王 欣
上海理工大学健康科学与工程学院 食品质量与安全研究所, 上海 200093
菲律宾蛤仔(Ruditapesphilippinarum)为一种海水贝类,属软体动物门、双壳纲、帘蛤目、帘蛤科,俗称花蛤。因其味道鲜美,营养丰富,低脂高蛋白,富含多种微量元素而深受消费者喜爱,是我国主要的海产经济贝类之一[1]。但受生长环境和滤食习惯的影响,鲜活菲律宾蛤仔中微生物数量较多,组成复杂,这导致菲律宾蛤仔在贮藏期间极易因微生物的生长及酶的作用而造成品质劣变。有调查对上海市生食贝类水产品安全性进行了半定量风险评估,显示贝类水产品中副溶血性弧菌平均检出率为43.7%,过量使用会危害人体健康[2]。
虽然低温冷藏是抑制微生物的生长及酶的作用,延长鲜活水产品货架期的有效方法之一[3-4]。但研究表明,鲜活水产品在低温保藏过程中仍会因微生物的生长而发生腐败变质。例如,李念等[5]的研究发现罗氏沼虾在冷藏过程中的菌落总数随时间延长而显著增加,冷藏3 d后即超过6 lg(CFU/g),使虾体发生腐败变质;
马锦涛等[6]的研究证明低温保藏减缓了金枪鱼片的腐败速度,但菌落总数仍逐渐增加,且在第6 d达到7 lg(CFU/g),使鱼片失去食用价值;
CHEN H B等[7]研究了不同产地的牡蛎腮中微生物数量与腐败的关系,发现其初始菌落总数为3.1~4.5 lg(CFU/g),而冷藏8 d后则显著上升至7.8~8.8 lg(CFU/g)到达货架期终点。因此,对鲜活水产品冷藏过程中的腐败微生物进行研究是提高其品质,延长货架期的重要基础。
另一方面,随着保藏时间的延长,鲜活水产品中的菌相也会发生变化,其中,一部分数量较多,具有最适生活力的微生物菌群在鲜活水产品的腐败变质过程中发挥着主要的作用,这部分微生物被称为该类产品的特定腐败菌(specific spoilage organism, SSO)[8]。例如,赵宏强等[9]对冷藏鲈鱼片的优势腐败菌进行了研究,发现冷藏鲈鱼的优势腐败菌主要为希瓦氏菌属与假单胞菌属;
而于淑池等[10]综合贮藏末期所占比例及致腐能力结果确定腐败希瓦氏菌为冷藏卵形鲳鲹的优势腐败菌;
CAO R等[11]研究发现太平洋牡蛎中的优势微生物为假单胞菌(22%)和弧菌(20%),且随着贮藏时间的增加,假单胞菌的比例明显增加;
MADIGAN TL等[12]则发现假单胞菌属和弧菌属在去壳牡蛎变质过程中起重要作用。因此,特定腐败菌是造成产品腐败的主要原因,对特定腐败菌的研究可以为后续进行针对性的抑菌保鲜提供参考依据。然而,文献调研表明,已有研究多围绕菲律宾蛤仔的加工保藏方法开展,而对其冷藏过程中的优势腐败菌的研究相对较少。
基于以上分析,本研究拟采用传统的培养方法重点对菲律宾蛤仔冷藏过程中的腐败菌进行分离及鉴定,并通过控制温度、pH、盐浓度和贝汁浓度来研究不同条件对腐败菌生长特性的影响,最后比较腐败因子进而判断致腐能力确定优势腐败菌,旨在为控制鲜活菲律宾蛤仔冷藏过程的品质劣变提供参考。
1.1 材料、试剂及仪器
鲜活菲律宾蛤仔,原产地辽宁省丹东市,购于上海壹佰米网络科技有限公司。低温盐水冷藏运输,半小时内送达实验室后置于冰箱冷藏(4 ℃)备用。
平板计数琼脂(PCA)、溶菌肉汤(LB)固体及液体培养基、革兰氏染液试剂盒、氧化酶试纸等均购自青岛海博生物技术有限公司;
氯化钠、氢氧化钠、硼酸、磷酸缓冲液、三氯乙酸、氧化镁等均为分析纯,购于国药集团化学试剂有限公司。
GHP-9080型隔水式恒温培养箱,上海一恒科学仪器有限公司;
YXQ-75G立式压力蒸汽灭菌锅,上海博迅实业有限公司医疗设备厂;
VD-850型桌上式洁净工作台,苏州沪净净化有限公司;
XW-80A微型旋涡混合仪,海门市其林贝尔仪器制造有限公司;
3-18R医用离心机,湖南恒诺仪器设备有限公司;
Nikon ECLIPSE E200MV生物显微镜,上海尼康仪器有限公司;
URT210-C多功能微孔板检测仪,美国伯腾仪器有限公司;
K1100全自动凯氏定氮仪,海能未来技术有限公司;
FP-1100-C生长曲线仪,美国赛默飞世尔科技有限公司。
1.2 试验方法
首先,对鲜活菲律宾蛤仔4 ℃冷藏过程的感官评分、菌落总数及挥发性盐基氮(total volatile base nitrogen, TVB-N)进行分析;
在对其中培养过程中占比较多,形态显著的菌株进行分离、纯化及鉴定的基础上,进一步研究不同因素对几种腐败菌的生长特性的影响,最后还分析了各菌株的致腐能力,从而确定冷藏鲜活菲律宾蛤仔中的优势腐败菌。
灭菌贝汁的制备: 参照宋绍华等和PARLAPANI F F等[13-14]的方法制备灭菌贝汁,将新鲜菲律宾蛤仔先用无菌水清洗、沥干后用洁净小刀去壳取贝肉。将贝肉绞碎后按肉∶水=1∶2(m/v)的比例加蒸馏水煮沸15 min,冷却后置于冰箱中静置过夜,以除去表层浮油;
8 000 r/min离心10 min后,取上清液121 ℃, 20 min灭菌,冷至室温后4℃保存备用。
1.3 分析方法
1.3.1菲律宾蛤仔感官评分
由12名实验室人员组成感官评定小组,依据表1综合评定样品的感官得分。

表1 菲律宾蛤仔感官评分标准评价表
1.3.2菌落总数及挥发性盐基氮(TVB-N)测定
参照GB 4789.2—2016《食品微生物学检验:菌落总数测定》进行菌落总数的测定[15]。参照GB 5009.228—2016《食品安全国家标准 食品中挥发性盐基氮的测定》进行TVB-N的检测[16]。
1.3.3腐败菌的分离及鉴定
在冷藏过程中,每隔1 d选取菌落数在30~300的平板,挑取其中占比较多,形态显著的菌落进行平板划线分离及纯化传代培养,并进行菌落形态革兰氏染色、氧化酶及过氧化氢酶试验。在此基础上,参照郑瑞生等[17]对即食鲍鱼腐败菌鉴定的方法,进行分子生物学鉴定[18],具体步骤如下:
菌落PCR:先取2 mL离心管,加10 μL(双蒸水)ddH2O,再用枪头挑起一个菌落加入其中,吹吸混匀,待用。PCR 扩增反应体系为模板(菌液)1.0 μL、ddH2O 10.5 μL、引物27F 0.5 μL、引物1492R 0.5 μL、PSIS 12.5 μL,将其振荡混合均匀后上机。PCR扩增条件:95 ℃预变性3 min后,98 ℃持续变性10 s,55 ℃退火30 s,72 ℃延伸90 s,共30个循环,最后72 ℃延伸10 min。随后,取PCR 扩增产物进行琼脂糖凝胶电泳并将纯化后的16S rDNA序列送上海华大基因有限公司进行测序,登陆NCBI网站进行同源性分析。
1.3.4不同因素对腐败菌生长的影响
调整菌液浓度为107CFU/mL,选择LB液体培养基,研究温度、pH、NaCl浓度及添加贝汁对各腐败菌株的生长状况的影响。具体如下:
(1) 温度:
在无菌96孔板中分别加入20 μL不同腐败菌的菌液,再加入180 μL灭菌 LB液体培养基后,分别置于25 ℃、30 ℃和37 ℃下培养,测定培养初始(0 h)和24 h时的OD600 nm,计算各组的ΔOD600 nm值 (ΔOD600 nm=OD600 nm(培养24 h)-OD600 nm(初始)),分析培养温度对各腐败菌株生长的影响[19]。
(2) pH:
LB液体培养基灭菌前用0.1 mol/L NaOH和0.1 mol/L HCl调pH分别至6.0、7.0、8.0,孔板装样比例及量不变,培养温度为37 ℃,测定培养初始(0 h)和24 h时的OD600 nm,计算各组的ΔOD600 nm值,分析体系的pH对各腐败菌株生长的影响。
(3) NaCl浓度:
使LB液体培养基中的NaCl浓度分别为1%、2%、3%和6%,装样比例及量不变, 培养温度为37 ℃,测定培养初始(0 h)和24 h时的OD600 nm,计算各组的ΔOD600 nm值,分析NaCl浓度对各腐败菌株生长的影响。
(4) 贝汁浓度
使LB液体培养基中的灭菌贝汁浓度分别为0%、5%、10%,装样比例及量不变,利用FP-1100-C全自动生长曲线仪检测各菌株24 h的生长情况,进而将收集到的数据通过Com Base网站进行生长动力学分析。所得到的生长动力学参数主要有菌液样本的初始浓度lg值(lgN0)、 37 ℃下培养24 h后菌液样本的末期浓度lg值(lgNmax)、 生长延滞期(Lag/h)、 最大比生长速率(μmax/ h-1),用回归系数(R2,越接近1,模型拟合度越高)、均方误差(MSE)对模型的准确性进行评价。
1.3.5不同菌株腐败能力的比较
调整菌液浓度为107CFU/mL,各取0.5 mL菌液与50 mL无菌贝汁混匀后, 4 ℃冷藏保存,分别在24 h和72 h时取样液进行菌落总数和TVB-N的测定,参照许振伟等[20]的方法,以腐败代谢产物产量因子(YTVB-N/CFU)为评价指标,对7株菌的腐败能力进行定量分析,YTVB-N/CFU的计算公式如下:
(1)
式中:(TVB-N)0、(TVB-N)s分别表示24 h和72 h的TVB-N含量,mg/100 mL,N0、Ns分别表示24 h和72 h的菌落总数,CFU/g。
1.4 数据处理
试验均为三次平行三次重复,采用 Microsoft Excel 2019 和 SPSS 26.0 对数据进行分析,采用 ANOVA 进行组间差异显著性分析(p<0.05)。采用 Origin2018 绘图。
2.1 菲律宾蛤仔冷藏过程中菌落总数,感官评分及TVB-N的变化
2.1.1菌落总数及感官评分的变化
微生物的生长是导致鲜活水产品腐败的主要原因,一般可用菌落总数反映其受微生物污染的程度。感官评分则可以反映食品的品质优劣及其可食用性。菲律宾蛤仔冷藏过程中菌落总数和感官评分的变化如图1所示。

图1 菲律宾蛤仔冷藏过程中菌落总数和感官评分的变化
图1表明,随冷藏时间的延长,菲律宾蛤仔的菌落总数呈上升趋势,冷藏1 d后,菌落总数从最初的4.81 lg(CFU/g) 显著增加至5.01 lg(CFU/g)(p<0.05),这已超出了国际微生物标准委员会关于新鲜双壳类软体动物的微生物食用限量标准(105CFU/g)[21]。且冷藏3 d后菌落总数进一步增加至5.53 lg(CFU/g)(p<0.05),说明腐败程度加剧,潘澜澜等[22]发现,冷藏4 d后死亡的海湾扇贝的菌落总数有相似的结果5.6 lg (CFU/g)。伴随着菌落总数的增加,感官评分则出现了明显的降低,如图1中所示,新鲜贝肉色泽相对明亮且肉质紧实,感官评分为9.2分,随冷藏时间的延长,贝肉的光泽及弹性降低,冷藏3 d后感官评分已低至5.9分(p<0.05),且有异味产生;
冷藏4 d后样本全部死亡,肉质泛白软烂,失去弹性,异味重。这是由于,冷藏过程中微生物的生长代谢加速了营养物质的降解, 致使菲律宾蛤仔逐渐出现肉质松软、色泽暗淡、散发腥臭味等腐败现象, 感官品质均呈下降趋势[23]。
2.1.2TVB-N的变化
挥发性盐基氮(TVB-N)是样品中的蛋白质在微生物和内源酶的作用下分解生成的氨类和胺等挥发性碱性含氮物质,是评价水产品新鲜度的重要指标,可以反映腐败的程度。菲律宾蛤仔冷藏过程中TVB-N的变化如图2所示。

图2 菲律宾蛤仔冷藏过程中TVB-N的变化
通常情况下,TVB-N与微生物的生长有一定的相关性。从图2可以看出,与图1中菌落总数的变化类似,菲律宾蛤仔的TVB-N值随冷藏时间的延长而增大,冷藏1 d后,其TVB-N值已显著增大至12.23 mg/100 g,而冷藏3 d后则进一步增大至18.27 mg/100 g,超出了国内农业部关于海水贝类(NY/T 1329—2017绿色食品 海水贝)的限值(15 mg/100 g),这也与图1中感官评分的显著降低相符,说明样品已出现了明显的腐败变质。
2.2 腐败菌株的分离鉴定
对菲律宾蛤仔冷藏过程中占比较大、形态显著的菌落进行分离纯化。最终根据菌落形态和显微镜检结果将得到的菌株归为7类,具体的菌株特征如表2所示。

表2 冷藏菲律宾蛤仔分离出的优势菌株的特征比较
由表2可知, 除1号菌株为革兰氏阳性球菌外,其余6株均为革兰氏阴性杆菌。2号、3号、5号、6号、7号菌氧化酶试验呈阳性,说明这几株菌为不发酵的革兰氏阴性杆菌。过氧化氢酶试验中,仅4号菌株呈阴性,而其余菌株均呈阳性,说明这些菌株中均存在过氧化氢酶。进一步对7株细菌进行16S rRNA鉴定,将基因序列与NCBI中的基因总库进行比对,选取其中最相似的序列进行相似性分析,结果如表3所示。

表3 各菌株的16S rRNA鉴定结果
从表3可以看出,7株菌的16S rDNA 序列与模式菌株序列的相似度均大于98%,再结合表2所得的形态颜色,生理生化结果,与表3各株细菌种类的形态特征基本吻合,说明鉴定结果可靠。1号、4号菌株为不动杆菌属,2号菌株为希瓦氏菌属,3号、6号及7号菌株为假单胞菌属,而5号菌株为金黄杆菌属,这与ODEYEMI O A等[24]的研究结果相似,他们从4 ℃冷藏贻贝中也主要分离得到了不动杆菌,希瓦氏菌及假单胞菌。将上述7株菌分别重新接种到新鲜菲律宾蛤仔样品中,在4 ℃冷藏条件下,发现都加快了样品的腐败速度,冷藏菲律宾蛤仔提前出现色泽变暗,肉质软烂以及腐臭异味等腐败现象,也能说明分离出的7株菌是冷藏菲律宾蛤仔的腐败菌。
2.3 不同因素对腐败菌生长的影响
进一步分析7株腐败菌在不同温度、pH、NaCl及贝汁浓度下的生长特性。
2.3.1温度
温度对微生物的存活及生长有重要影响。7株腐败菌在不同培养温度下的生长状况如图3所示。

图3 7株腐败菌在不同温度下的生长状况
从图3可以看出,培养温度对7株腐败菌的生长状况有一定影响。整体来看,不动杆菌属(1号及4号菌)生长情况相对较优,其次为可萨假单胞菌(3号菌),而2号(腐败希瓦氏菌)和5号(金黄杆菌)生长情况相似。1号、4号、5号菌在30 ℃生长最佳,其ΔOD600分别为0.76、0.84、0.55;
3号、6号、7号菌在37 ℃生长最佳,其ΔOD600分别为0.67、0.55、0.54,而2号菌在25 ℃生长最佳,ΔOD600为0.54,显著高于其他两个温度下的结果(p<0.05),朱彦祺等[25]也发现分离自大黄鱼的腐败希瓦氏菌在25 ℃的延滞期显著较短,而比生长速率较高。
2.3.2pH
7株腐败菌在不同pH环境中的生长状况如图4所示。

图4 7株腐败菌在在不同pH环境中的生长状况
2.3.3NaCl浓度
图4表明,培养环境的pH条件会对7株腐败菌的生长有一定的影响。除2号菌(腐败希瓦氏菌)外,其余6株菌在pH=7的中性条件下生长良好。4号菌(溶血不动杆菌)的生长受环境pH的影响较小,其ΔOD600均相对较高。2号菌(腐败希瓦氏菌)在pH=7~8时的生长量显著高于pH=6时的结果,这与YOON J H等[26]对分离自海水中的希瓦氏菌的最适生长pH的结果相符。
NaCl浓度对7株腐败菌生长状况的影响如图5所示。

图5 7株腐败菌在不同NaCl浓度下的生长状况
从图5可以看出,除2号菌外,添加1% 的NaCl有助于6株腐败菌的生长,其ΔOD600显著高于其他环境,但随着NaCl浓度的升高,各菌的ΔOD600又相对降低,说明菌的生长受到一定的抑制。其中,在1%NaCl下,4号菌(溶血不动杆菌)与1号菌(威尼斯不动杆菌)的ΔOD600高达0.78与0.57,生长相对旺盛;
而2号菌(腐败希瓦氏菌)的ΔOD600仅为0.31,生长相对缓慢,这与赵二科等[27]研究中发现的希瓦氏菌的ΔOD600值(0.2~0.4)类似。
2.3.4贝汁浓度
表4给出了7株腐败菌在不同贝汁浓度下的生长动力学参数信息。其中包括菌液样本的初始浓度lg值(lgN0)、37 ℃下24 h后菌液样本的末期浓度lg值(lgNmax)、生长延滞期Lag、最大比生长速率μmax以及对模型准确性的评价R2回归系数(越接近1,模型拟合度越高)、MSE(均方误差)。
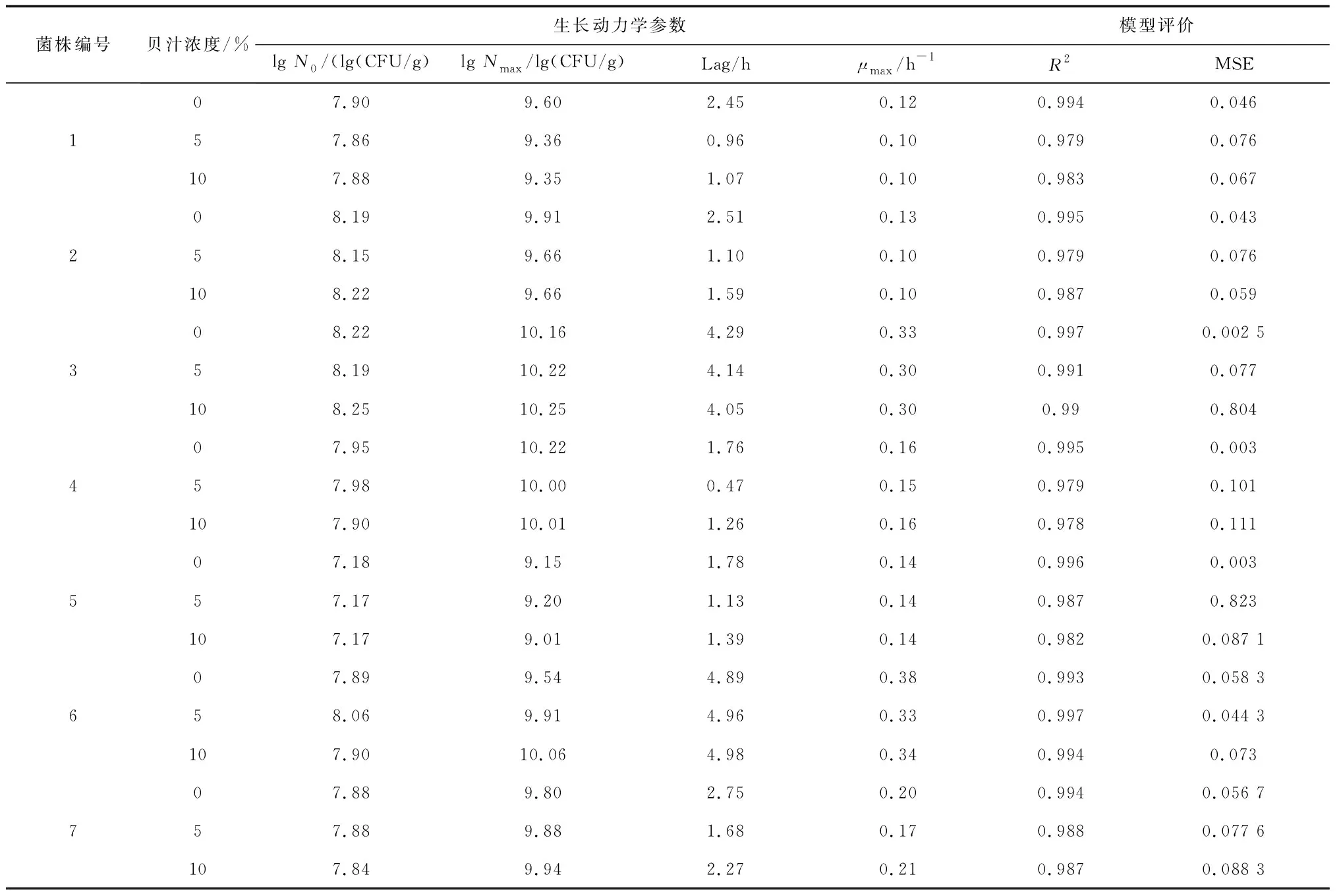
表4 贝汁浓度对7株腐败菌生长情况的影响
由表4可知,各模型的R2均大于0.93,且生长曲线都呈现出典型的“S”型特征,说明所采用的Baranyi and Roberts模型能准确描述7株腐败菌在不同贝汁浓度下的生长规律。贝汁浓度对各菌株的延滞期影响较大,除6号菌外,其余菌株的延滞期在添加贝汁后菌有明显的缩短,且与添加10%贝汁的结果相比,1号、2号、4号、5号、7号菌的延滞期在5%贝汁条件下更短,说明添加5%的贝汁可促进其生长繁殖,提高菌量。3号菌则在添加10%贝汁的条件下具有更短的延滞期。相比之下,添加贝汁对各菌株的比生长速率影响相对较小,说明贝汁环境不能加快各腐败菌的生长速度。Baranyi and Roberts模型拟合结果说明延滞期与贮藏贝汁浓度有一定的线性关系[28]。
2.4 不同菌株腐败能力的比较
将7株腐败菌分别接种至灭菌贝汁并冷藏24 h和72 h后测得的菌落总数、TVB-N以及由其计算得到的腐败代谢产物产量因子(YTVB-N/CFU)的结果如表5所示。

表5 分别接种7株菌后灭菌贝汁冷藏过程中的菌落总数,TVB-N及腐败因子
从表5可以看出,接种2(腐败希瓦氏菌)、3(可萨假单胞菌)、7(吲哚氧基假单胞菌)号菌株的贝汁并冷藏24 h后,其菌落总数显著较高(p<0.05),分别为4.01 lg(CFU/g)、3.98 lg(CFU/g)、3.89 lg(CFU/g),且接种了2(腐败希瓦氏菌)、3(可萨假单胞菌)号菌的贝汁的TVB-N值也显著较高(p<0.05),分别为2.03 mg/100 mL、1.96 mg/100 mL。而冷藏72 h后,接种1(威尼斯不动杆菌)、4(溶血不动杆菌)、5(金黄杆菌)、6(门多萨假单胞菌)号菌的贝汁的菌落总数显著较高(p<0.05),分别为6.29 lg(CFU/g)、6.26 lg(CFU/g)、6.13 lg(CFU/g)、6.24 lg(CFU/g)。但此时TVB-N值较高的样本是接种了2(腐败希瓦氏菌)、 3(可萨假单胞菌)和7(吲哚氧基假单胞菌)号菌的贝汁,分别为9.03 mg/100 mL、7.68 mg/100 mL和7.31 mg/100 mL。
Y(TVB-N/CFU)可以定量表征各腐败菌的腐败能力[9]。由表5可知, 2(腐败希瓦氏菌),3(可萨假单胞菌)及7(吲哚氧基假单胞菌)号菌的Y(TVB-N/CFU)值显著高于其他腐败菌(p<0.05),其中, 2号及7号菌的Y(TVB-N/CFU)高达1.30×10-5mg/CFU,1.28×10-5mg/CFU,而3号菌的Y(TVB-N/CFU)也高达9.69×10-6mg/CFU。说明腐败希瓦氏菌、吲哚氧基假单胞菌及可萨假单胞菌的致腐能力显著较高,可以判定为冷藏菲律宾蛤仔中的优势腐败菌。这与郝若伊等[29]在4 ℃冷藏鲍鱼中确定的优势腐败菌(腐败希瓦氏和假单胞菌)相似。
本研究在对冷藏菲律宾蛤仔的感官评分、菌落总数及挥发性盐基氮变化规律研究的基础上,采用传统的培养方法对其中的腐败菌进行分离和鉴定,研究了几种因素(温度、pH、盐浓度及贝汁浓度)对腐败菌生长特性的影响,最后通过比较腐败能力确定了优势腐败菌。结果表明,菲律宾蛤仔冷藏过程中菌落总数及TVB-N显著增大而感官评分相对减小。冷藏3 d后失去食用价值。分离纯化后获得7株腐败菌,经过形态、生理生化及16S rRNA 序列分析后确定其分别归属不动杆菌(威尼斯不动杆菌;
溶血不动杆菌)、假单胞菌(可萨假单胞菌;
门多萨假单胞菌;
吲哚氧基假单胞菌)、希瓦氏菌(腐败希瓦氏菌)及金黄杆菌属。不动杆菌属的生长受温度的影响较小,而腐败希瓦氏菌在较低温度(25 ℃)的生长相对较好;
不动杆属类受环境pH影响最小,腐败希瓦氏菌在碱性环境中生长更好;
除腐败希瓦氏菌外,其余6株菌在中性及添加1% NaCl的环境中生长良好。添加5%的贝汁有助于缩短1、2、4、5、7号菌株的生长延滞期,促进其生长繁殖,但对比生长速率影响较小。腐败希瓦氏菌、吲哚氧基假单胞菌、可萨假单胞菌的腐败因子显著较高,说明这3株菌为冷藏菲律宾蛤仔的优势腐败菌,且致腐能力:腐败希瓦氏菌>吲哚氧基假单胞菌>可萨假单胞菌。该研究为后续靶向调控鲜活菲律宾蛤仔冷藏过程的品质劣变,延长货架期可提供一定的参考。
下一篇:45钢厚板火切开裂成因分析